
Ingrese su ID de correo electrónico
EJE HORMONAL SEXUAL FEMENINO. CARACTERÍSTICAS GENERALES.
El eje endocrino (figura 1) que regula las hormonas sexuales femeninas comienza en el hipotálamo con la secreción hipotalámica de Kisspeptina [1]Las Kisspeptinas, neuropéptidos del grupo RFamida, son una familia de péptidos codificados por el gen Kiss1, localizado en el brazo largo del cromosoma 1 (1q32), que actúan uniéndose al receptor acoplado a la proteína G denominado GPR54 o Kiss1R.La estimulación de GPR54 por la Kisspeptina constituye el más potente estímulo fisiológico para la secreción de GnRH y es indispensable para alcanzar los elevados niveles de éste necesarios para … Continúe leyendo, hormona que participa de forma importante en el inicio de la pubertad y el ciclo ovárico, que a su vez regula la secreción de la GnRH o LHRH (o luteoliberina) y el GAP (péptido asociado a la GnRH )(fragmento de ésta (1-13)), encargados de regular la secreción de las gonadotropinas adenohipofisarias LH y FSH, las cuales regulan a su vez la secreción de los estrógenos, andrógenos y progesterona de origen ovárico [2]Los mensajeros químicos utilizados en el eje reproductor femenino son los mismos que los utilizados por el eje reproductor masculino. Varían las concentraciones plasmáticas y los patrones de retroalimentación que gobiernan el funcionamiento del eje.

Figura 1: Eje hormonal sexual femenino.
La regulación que a partir de la pubertad muestra un patrón cíclico que da lugar al ciclo menstrual o ciclo ovulatorio, con base en un cambio del patrón secretor de las gonadotropinas. Lo que supone que la capacidad reproductora de la mujer no sea continua, diferenciándose del hombre el cual produce unos dos millones de células germinales frente a una, por regla general, que produce la mujer cada 28 días. Este ciclo se interrumpe solo si hay fertilización del óvulo, o cuando cesa de forma natural (menopausia).
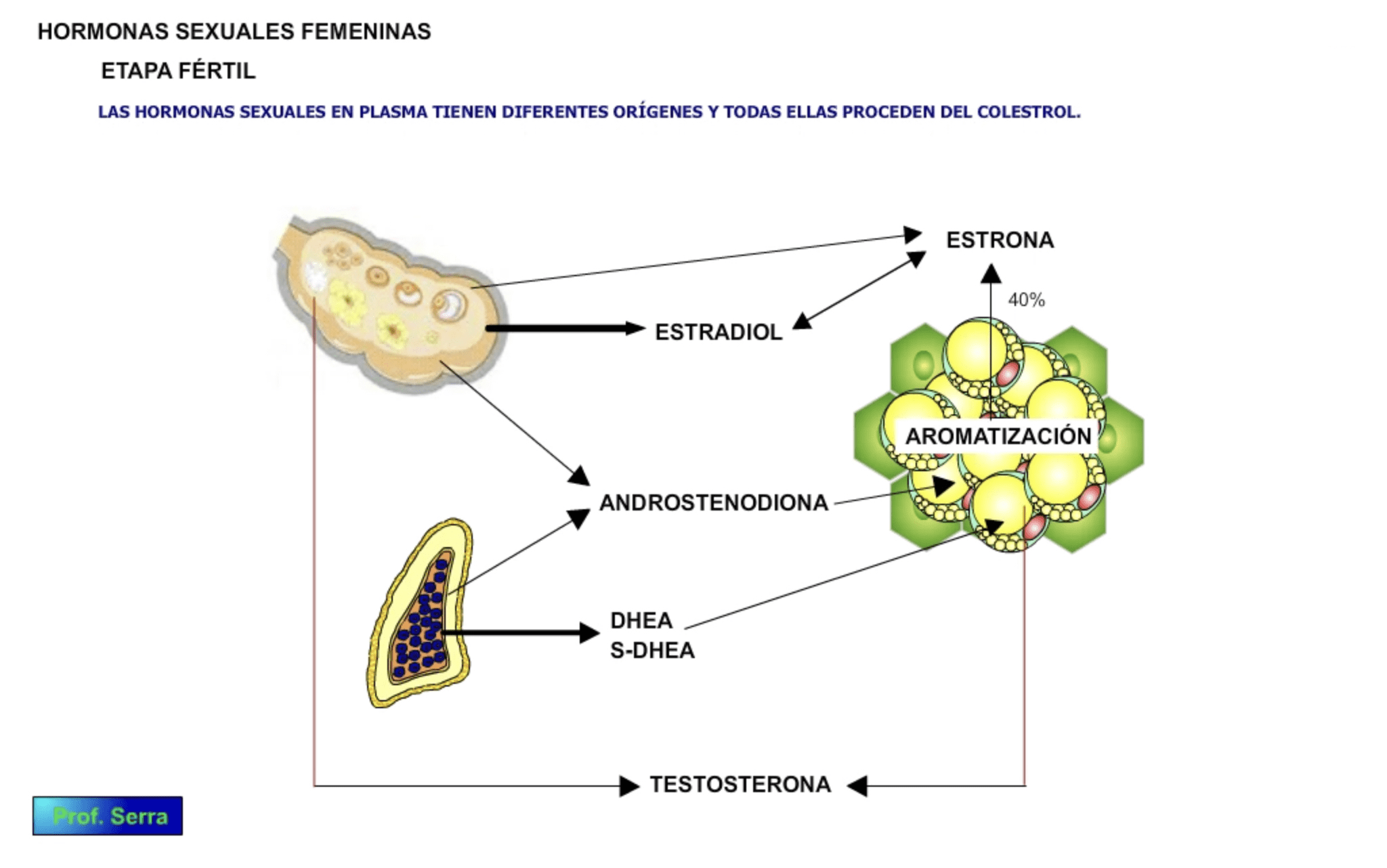 No todas las hormonas sexuales en la mujer son de origen ovárico (figura 2), sino que una parte procede de la capa reticular de las suprarrenales y del tejido adiposo. Fracción esta última que adquiere importancia a partir de la etapa posmenopáusica, donde el ovario disminuye el nivel de secreción de hormonas sexuales.
No todas las hormonas sexuales en la mujer son de origen ovárico (figura 2), sino que una parte procede de la capa reticular de las suprarrenales y del tejido adiposo. Fracción esta última que adquiere importancia a partir de la etapa posmenopáusica, donde el ovario disminuye el nivel de secreción de hormonas sexuales.

Por otra parte, este eje está sometido a la acción indirecta de otros factores que pueden alterar su normal funcionamiento como: alteraciones hormonales, infecciones, desnutrición y factores sicológicos.

Figura 2: Orígenes de las hormonas sexuales femeninas según etapas sexuales de la mujer.
GÓNADAS FEMENINAS
El ovario (figura 3), como órgano endocrino, pertenece al eje endocrino sexual femenino. Es una estructura par unida al cuerpo del útero mediante el ligamento ovárico y conectado a la cavidad uterina mediante el oviducto o trompa de Falopio (de 10 a 15 cm ), con paredes finas y recubiertas por proyecciones ciliadas densas (fimbrias) que recepcionan el óvulo liberado. Es en la zona intermedia del conducto (la ampolla) donde se produce la fertilización del óvulo. Este óvulo pasa al útero a través del istmo, última parte del conducto que actúa como esfínter. El movimiento del óvulo se facilita por las contracciones del músculo liso, los cilios y las secreciones. En la mujer adulta cada ovario pesa de 6 a 10 gramos.

Figura 3: Morfología de los ovarios.
El útero es un órgano hueco diseñado para el crecimiento del feto en caso de que se produzca la fertilización. Con una alta capacidad de crecimiento, desde el tamaño de un puño en la mujer no embarazada hasta el tamaño de un niño de unos tres kilos. Tiene dos tipos de tejidos: el miometrio formado por varias capas de músculo liso, y la más interna el endometrio, formado por una capa de estroma próxima al miometrio y una capa epitelial superficial, interrumpida por las glándulas uterinas que penetra la capa de estroma y posee células secretoras columnares.
El cérvix une la vagina con el útero y sirve como barrera mecánica a los espermatozoides, y protege contra la infección.
La vagina (conducto muscular elástico) posee un epitelio mucoso poliestratificado que secreta un moco rico en glucógeno cuya fermentación producida por el bacilo de Döderlein le da carácter ácido a la secreción vaginal. Esta secreción varía durante el ciclo y sirve de lubrificante durante el coito.
El ovario maduro está formado por una corteza y una médula o hilio por donde entran y salen los vasos sanguíneos, linfáticos y nervios.
En la corteza [3]La corteza ovárica está formada por dos capas de fuera hacia dentro: el estroma (capa de sostén): con una capa epitelial externa denominada capa germinal, nombre que puede dar una falsa idea, pues se sabe que los óvulos proceden del endodermo y emigran a los ovarios durante el desarrollo embrionario, seguida por otra capa denominada falsa capa albugínea, con fibroblastos; y una más interior: el estroma propiamente dicho, con tejido conjuntivo, … Continúe leyendo encontramos células de sostén, fibroblastos, folículos en diferentes estados de maduración, los cuerpos lúteos, el cuerpo albicans y las células intersticiales, tecales y deciduales. Las tecales migran hacia la superficie folicular, y las intersticiales secundarias [4]Las células intersticiales primarias aparecen en la semana nº 12 del feto y desaparecen a las ocho semanas. que proceden de la región tecal una vez que el folículo se vuelve atrésico.
También tenemos las células intersticiales del hilio que secretan andrógenos.
El cuerpo lúteo también presenta células derivadas de la transformación del folículo de Graaf que ha ovulado y son secretoras de progesterona y otras hormonas.
OVOGÉNESIS
Los folículos son la unidad funcional del ovario, ya que al final de su proceso de maduración resulta la liberación de un óvulo.
El ovario adulto es distinto al preadulto por los diferentes estadios foliculares que pueden observarse. Por tanto distinguiremos dos etapas: la preadulta que se inicia en el ovario del feto, en las primeras semanas del embarazo materno y la adulta, que comienza con el primer ciclo menstrual del sujeto adulto.

Figura 4: Ovario prepuberal.
OVARIO PREADULTO: Las células germinales primarias u ovogonias se originan en el endodermo del embrión y durante las 3-5 semanas primeras emigran a las crestas genitales (figura 5). Unas 2000 ovogonias invaden la corteza ovárica e inician su proliferación mediante mitosis. Cuando llegan a unas 600.000 (hacia la semana 8), algunas cesan la mitosis e inician la primera división meiótica, otras continúan proliferando y a los 5 o 6 meses alcanzan unos 7×106 ovogonias. En este momento todas entran en meiosis parándose en la profase de la primera división meiótica y así se quedan hasta los 50 años. Estas ovogonias se rodean de células ahusadas y se separan del estroma mediante una membrana basal, dando lugar a los folículos primordiales. Un gran número de ellos entran en regresión (atresia).
A partir del 6º mes de gestación (figura 6) estos folículos primordiales maduran proliferando las células ahusadas para formar una sola capa de células granulosas cuboidales, rodeadas por la membrana basal. Estas células segregan mucopolisacáridos, dando lugar a la formación de un halo traslúcido "zona pelúcida " que rodea al oocito. Se forma así el folículo primario. Esta transformación se realiza de forma independiente a las hormonas periféricas y dentro de un ambiente regulador propio.
Tras el nacimiento y durante la etapa prepuberal se observa esta transformación continua de folículos primordiales en folículos primarios y así siguen durante toda la vida fértil de la mujer.
Al nacer el número de folículos primordiales ha disminuido considerablemente, quedando en unos 2×106, siguiendo el proceso de regresión (atresia) durante toda la vida fértil de la mujer, ya que sólo se usan unos 500 en toda esa etapa. Cuando comienza la pubertad solo quedan unos 300.000.
 Figura 5: Ovario prepuberal: Fase 1: Primeras semanas .
Figura 5: Ovario prepuberal: Fase 1: Primeras semanas .

Figura 6: Ovario prepuberal. Fase 1: 6 meses de gestación.

Figura 7: Ovario prepuberal: Fase 1: Antes de la pubertad.
OVARIO ADULTO: Una vez iniciado el ciclo fértil y con la aparición de las gonadotropinas, en cada ciclo se activan varios folículos primarios que son estimulados para su transformación, proceso que supone una tres semanas. Las células granulosas de estos folículos activados se dividen formando varias capas y a su vez, las células intersticiales se aproximan a la lámina basal y se transforman en células tecales (teca interna y externa) secretoras de andrógenos bajo el estímulo de la LH. Así se transforman en folículos preantrales o secundarios.
Los folículos se van agrandando y muchos de ellos degeneran en este proceso (folículos atrésicos). Sus células se hacen más sensibles a la LH y FSH y secretan bajo el estímulo de estas gonadotropinas, hormonas sexuales femeninas y masculinas. Las células granulosas comienzan a secretar diferentes productos y actúan como filtros del plasma, concentrando en el interior del folículo un gran número de sustancias y hormonas, dando lugar al antro y al folículo terciario ( 100 a 200 µm).
Figura 8: Ovario adulto. Segunda fase. Folículo creciente.
Los folículos siguen creciendo por acumulo de sustancias en el antro y el ovocito se rodea de una capa de células formando una especie de evaginación hacia el antro ("cúmulus oophurus"). Las células que rodean inmediatamente al ovocito se denominan corona radiante.

Figura 9: Ovario maduro. Tercera fase. Folículo de Graff.
Por mecanismos no bien conocidos, de todos los folículos seleccionados al final solo sigue uno y a veces dos, el resto degenera [5]En condiciones de estímulo prolongado de gonadotropinas pueden seleccionarse varios folículos.. Éste es el folículo de Graaf (figura 9) con un tamaño de 2 a 2,5 cm y una fuerte presión hidrostática antral. La atresia aparece por una disminución de la FSH plasmática debido a un aumento del estradiol (que inhibe la liberación de FSH). En el folículo seleccionado, la FSH se concentra en el antro y disminuye el número de receptores para dicha hormona. La alta concentración antral de estradiol amplifica el efecto mitogénico de la FSH y la actividad aromatasa. Por otro lado, el incremento de los niveles de andrógenos en forma no aromatizables (DHEA) en los folículos no seleccionados, antagoniza con la acción del estradiol y supone su regresión.
A medida que sigue creciendo el folículo seleccionado se va vascularizando, lo que favorece el aporte de colesterol y de gonadotropinas y la formación de estradiol. El pico de LH que se produce a mitad del ciclo determina cambios rápidos en el folículo seleccionado, que afectan a la división meiótica del ovocito, al tipo de secreción estrogénica, activación de enzimas proteolíticas, liberación de prostaglandinas, histamina, etc., así como un incremento importante del flujo sanguíneo que hace al folículo hiperémico y edematoso.
Tanto las células tecales como las granulosas producen activador del plasminógeno y plasmina, la cual degrada la pared folicular y favorece la destrucción del tejido conjuntivo de la matriz. La alta presión intrafolicular, la disgregación de la lámina basal y de las células foliculares y adelgazamiento de la pared ovárica en la región del folículo supone la rotura folicular y expulsión de la masa cúmulo-ovocito. Aquí también intervienen las prostaglandinas E y F que aumentan tras el pico LH. La LH también estimula la producción de progesterona que favorece la distensibilidad de la pared folicular y la activación de las enzimas proteolíticas. Justo antes de la ovulación se completa la 1ª división meiótica formándose el primer cuerpo polar que degenera y el ovocito secundario que recibe la mayor parte del citoplasma. Se produce la 2ª división meiótica y se detiene en la metafase hasta que llegue el espermatozoide. Es posible que el pico de LH inhiba o neutralice al factor inhibidor de la meiosis.

Figura 10: Ovario maduro. Ovulación.
En esta situación el ovocito es expulsado del folículo de Graaf (figura 10), pasando al oviducto donde sobrevive unas 72 horas. Si se produce fertilización se completa rápidamente la segunda meiosis, formándose un segundo cuerpo polar y un gran cigoto.
Del folículo de Graaf evacuado se forma el cuerpo lúteo. Las células granulosas y tecales se desorganizan y la pared folicular se colapsa y contornea. Los vasos lo invaden y el centro folicular se llena de sangre. Estas células se luteinizan (incluso antes de la expulsión del óvulo) y comienzan a producir progesterona, relaxina, oxitocina, esteroides sexuales, prostaglandinas, etc.
Todas estas secreciones sirven para mantener el embarazado durante los tres primeros meses, si éste se produce. Si no hay fertilización, a los 9 u 11 días el cuerpo lúteo degenera siendo remplazado por tejido fibroso, dando lugar al cuerpo albicans (figura 11). Si se produce fecundación, el cuerpo lúteo no degenera gracias a la acción de la HCG (Hormona gonocoriónica).

Figura 11: Ovario maduro. Fase lútea.
![]()
 Views Today : 189
Views Today : 189 Views This Month : 1820
Views This Month : 1820 Views This Year : 27443
Views This Year : 27443 Total views : 313506
Total views : 313506