
Ingrese su ID de correo electrónico
CATECOLAMINAS DE LA MÉDULA ADRENAL
Las catecolaminas circulantes proceden en su mayoría de la médula adrenal (figura 1)[1] Ensanchamiento y especialización de un ganglio simpático, cuyas neuronas postganglionares han perdido su axón y secretan su contenido directamente al torrente circulatorio., que las secreta tras el estímulo nervioso simpático adecuado.
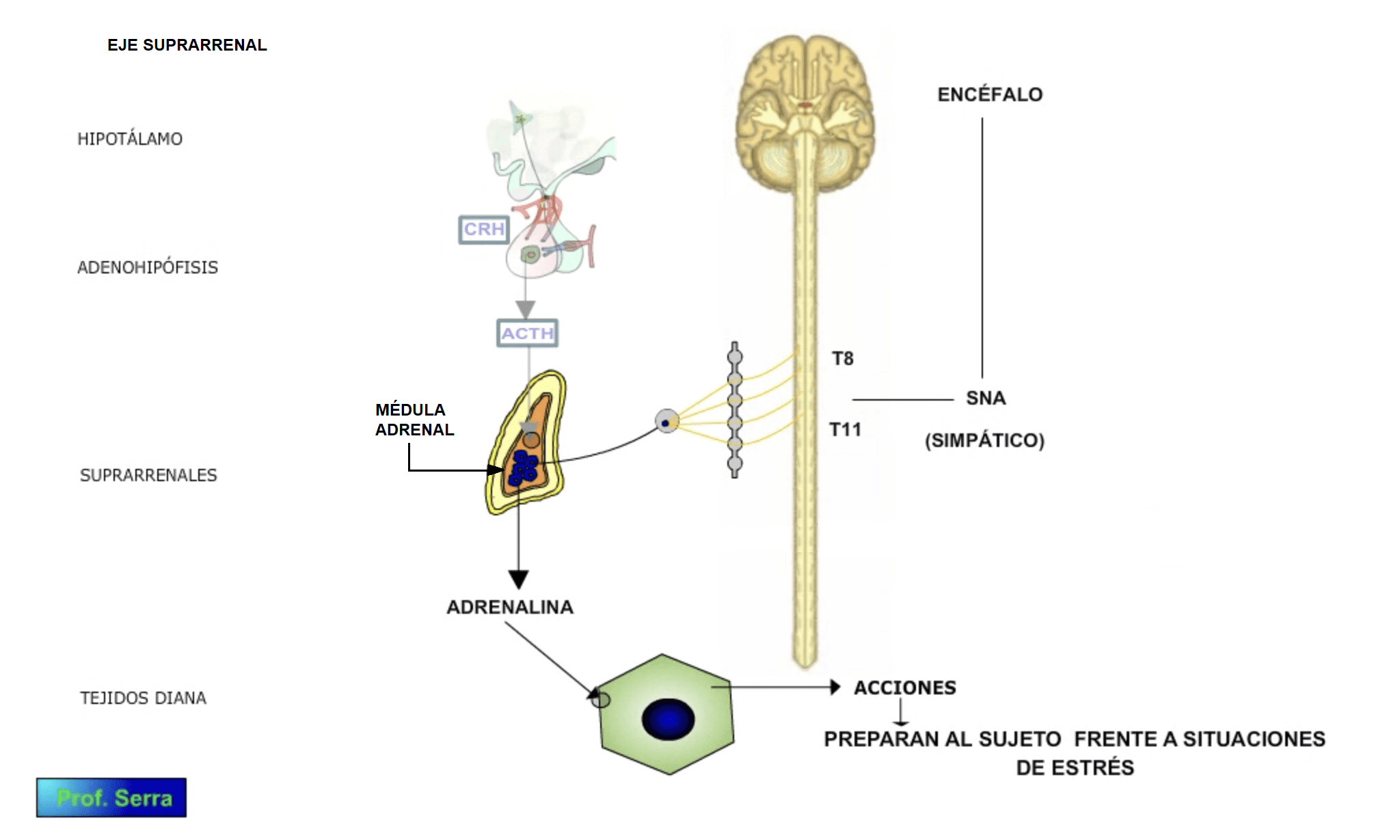
Figura 1: Eje suprarrenal-catecolaminas y sistema nervioso simpático.
Su origen es neuroectodérmico y a las siete semanas de gestación, las células de la cresta neural invaden el córtex adrenal fetal, dando lugar a la médula adrenal, que en el nacimiento es totalmente funcional.
Sobre las células medulares adrenales, las cuales están en estrecha relación con el sistema arteriovenoso del órgano, dejan sus terminaciones las fibras preganglionares simpáticas colinérgicas. Dentro de estas células, llamadas cromafines, se pueden distinguir numerosos gránulos de 100 a 300 nm de diámetro, los cuales contienen catecolaminas (20%), ATP (15%), proteínas (35%), lípidos (20%) y encefalinas. El 85% de las catecolaminas es adrenalina (A) y el resto noradrenalina (NA).
La médula adrenal está compuesta por agrupaciones de células cromafines dispuestas alrededor de los capilares. Su estrecha relación con la actividad del sistema simpático hace que funcionalmente se le denomine al conjunto sistema simpaticomedular.
Dado el diseño vascular de este órgano, la médula adrenal también puede estar bajo el influjo de las hormonas sintetizadas en el cortex suprarrenal.
En los seres humanos, la principal catecolamina secretada por la médula adrenal es la ADRENALINA.
El descubrimiento de esta hormona fue realizado independientemente por los químicos japoneses Jokichi Takamine (Yamashima, 2003;[2]Yamashima T (2003). "Jokichi Takamine (1854–1922), the samurai chemist, and his work on adrenalin". J Med Biogr 11 (2): 95–102. [Abstrac], 1999 [3]Bennett M (1999). "One hundred years of adrenaline: the discovery of autoreceptors". Clin Auton Res 9 (3): 145–59. y su asistente Keizo Uenaka en 1900 . Takamine la aisló en 1901 (Takamine, 1901[4]Takamine J (1901). The isolation of the active principle of the suprarenal gland. Great Britain: Cambridge University Press. pp. xxix-xxx.),aunque los primeros extractos adrenales conteniendo adrenalina fueron extraídos por Napoleon Cybulski. La síntesis de esta hormona fue realizada de forma independiente por Friedrich Stolz y Henry Drysdale Dakin , en 1904. (Bennet, 1999[5]Bennett M (1999). "One hundred years of adrenaline: the discovery of autoreceptors". Clin Auton Res 9 (3): 145–59.).
ADRENALINA. BIOSÍNTESIS
Las catecolaminas producidas en la médula adrenal son la adrenalina o epinefrina (sustituto sintético), la noradrenalina o norepinefrina (sustituto sintético) y la dopamina, siendo la primera la que tiene principal acción hormonal. (figura 2)

Figura 2: Catecolaminas adrenales.
La síntesis de las catecolaminas (figura 3) comienza con la captación a partir del torrente circulatorio de la tirosina, la cual por acción de la tirosina hidroxilasa (más O2 y y el cofactor tetrahidrobiopterina) se transforma en 3,4 dihidrofenilalanina (DOPA). El control de esta primera enzima determina el paso limitante de esta reacción . El siguiente paso es la conversión de la DOPA en dopamina por intermedio de una descarboxilasa la cual requiere como cofactor al piridoxal fosfato. Toda esta secuencia biosintética se realiza en el citoplasma, de aquí la dopamina pasa al gránulo de secreción (gránulo cromafin), donde la dopamina ß-hidroxilasa cataliza su conversión a noradrenalina en presencia de O2 y un donador de hidrógeno como el ácido ascórbico. En el 15% de los gránulos la secuencia biosintética acaba aquí. En los restantes la NA pasa al citoplasma nuevamente y ahí por intermedio de la feniletanolamina-N-metiltransferasa y el S-adenosilmetionina como donador del grupo metilo, se obtiene la adrenalina la cual vuelve al gránulo de secreción donde se almacena.

Figura 3: Biosíntesis de las catecolaminas en la médula adrenal.
Biosíntesis de otros compuestos de interés biológico en la médula adrenal:
Junto a las catecolaminas se encuentran otros compuestos, los cuales tienen papeles biológicos reconocidos, aunque de algunos de ellos no se sabe su función en la médula adrenal. Entre estos destacan las encefalinas, la serotonina, sustancia P, neurotensina, somatostatina, GABA y dopamina.
Las encefalinas, meta-encefalina (Tyr-Gly-Gly-Phe-Met) y la leu-encefalina (Tyr-Gly-Gly-Phe-Leu) se encuentran en alta concentración en las células cromafines de la médula y parecen proceder de una proencefalina que contiene ambas secuencias. Estas encefalinas son almacenadas dentro de las vesículas cromafines y son co-secretadas junto con las catecolaminas y otros compuestos solubles.
La serotonina también se encuentra en las células cromafines de la médula aunque la fuente de la misma no se ha establecido aún, no obstante se encuentra tanto la serotonina como su precursor el 5′-hidroxitriptófano.
MECANISMO DE ACCIÓN
El principal efector medulo-adrenal es la adrenalina, ya que la noradrenalina requiere incrementar 10 veces su valor plasmático para evidenciar efectos fisiológicos. La mayoría de la noradrenalina plasmática procede por difusión de las terminales postganglionares simpáticas, mientras que toda la adrenalina procede de la médula adrenal, ya que no puede pasar la barrera hemato-encefálica.

Figura 4: Mecanismo de acción de las catecolaminas.
La mayoría de los tejidos presentan receptores para las catecolaminas medulo-adrenales, las cuales actúan a través de los receptores a y ß, los cuales muestran distinta capacidad de reacción a estas catecolaminas como se muestra en la tabla de esta figura 5.

Figura 5: Mecanismo de acción de las catecolaminas según el receptor activado.
El mecanismo de acción depende del tipo de receptor activado. Así los beta y alfa 2 actúan sobre el AMPc, los primeros incrementando su concentración y el segundo disminuyéndola como se puede ver en la figura. Los beta actúan sobre la subunidad Ns y los alfa 2 sobre la subunidad Ni.
Por otra parte los receptores alfa 1 actúan incrementando la entrada de calcio a través de la activación de canales de calcio, y a su vez activando a la fosfolipasa C con el consiguiente incremento de IP 3 y la actividad de la PKC.
ACCIONES DE LAS CATECOLAMINAS
Estas hormonas están diseñadas para preparar al individuo ante situaciones de emergencia (estrés), lo cual supone modificaciones de la glucemia (incremento de la glucosa en sangre), modificaciones cardiocirculatorias, respiratorias y modificaciones en la secreción hormonal de otros ejes endocrinos. En realidad son responsables de los ajustes homeostáticos rápidos y de corto plazo. Las acciones que vamos a ver corresponden tanto a la adrenalina como a la noradrenalina, aunque esta última requiere niveles en plasma muy altos para realizar dichas funciones.

Figura 6: Acciones de las catecolaminas en el metabolismo.
Las modificaciones en la glucemia suponen modificaciones en el metabolismo que afectan a:
– la captación de glucosa por los tejidos
– la movilización de sustratos energéticos
– la producción de glucosa por el hígado
– la movilización de hormonas que afectan a la glucemia
En el esquema de la figura 6, se pueden observar las acciones de la adrenalina en los distintos tejidos implicados en la glucemia:
– Se produce una disminución en la captación de glucosa por parte de los tejidos que la utilizan, excepto el cerebro. Disminuyendo la liberación de insulina y la acción de ésta sobre sus tejidos diana.
– En el músculo esquelético estimula la glucogenólisis vía un mecanismo dependiente del AMPc que activa a la fosforilasa. Pero como el músculo no tiene glucosa 6P posiblemente la glucosa no pase a la circulación y sea metabolizada formándose lactato e hiperlactemia característica. La liberación de este ácido láctico es utilizada por el hígado para producir más glucosa.
– En el tejido adiposo blanco actúa como agente lipolítico, por un proceso dependiente de AMPc que activa a la lipasa, dando lugar a la producción de ácidos grasos libres y glicerol que pasan a la circulación.
– En el hígado activa la glucogenólisis y la gluconeogénesis e inhibe la glucogenosíntesis. Esta acción la realiza a través de fosforilaciones dependientes del AMPc en su mayor parte, y por otros mecanismos no dependientes del AMPc (posiblemente el calcio iónico libre).
– En el páncreas endocrino, inhibiendo la liberación de insulina y estimulando la del glucagón.
– En el tejido adiposo pardo, incrementa la producción de calor con el consecuente incremento de la temperatura corporal.
Regula la [K + ] plasmático por cuanto éste disminuye al principio por su entrada en músculo esquelético, pero después se incrementa en plasma por su liberación hepática.

Figura 7: Acciones de las catecolaminas en el sistema cardiovascular.
Las acciones cardiovasculares (figura 7) las realiza incrementando el gasto cardiaco . Esta acción está mediada por receptores ß1 y a través de ellos se consigue: estimular la tasa de descarga del nodo sino atrial; la velocidad de conducción y la fuerza de contracción en el músculo atrial y ventricular.
La noradrenalina produce vasoconstricción en prácticamente todos los sitios, mientras que la adrenalina produce vasodilatación en músculo esquelético e hígado (ß2). El resultado final es una disminución de la resistencia total periférica. La noradrenalina aumenta la presión sistólica y la diastólica y por tanto la presión media. La adrenalina aumenta la sistólica y disminuye la diastólica dejando prácticamente igual la presión media o ligeramente aumentada. Ambas reducen el flujo sanguíneo en la piel. En el riñón se produce reducción de flujo sanguíneo por ambas, las cuales incrementan el flujo sanguíneo coronario.
El efecto final de estas acciones es incrementar el flujo sanguíneo y derivarlo hacia los músculos, hígado, corazón y cerebro.

Figura 8: Acciones de las catecolaminas en el músculo liso.
Las acciones de ambas catecolaminas sobre el músculo liso visceral (figura 8) son variables y dependen del tipo de receptor que éstos presenten. Así la relajación de este músculo se consigue mediante la activación de receptores ß2 , mientras que la contracción se realiza por receptores α1.
El músculo liso gastrointestinal se relaja con la adrenalina reduciendo ésta el tono y la amplitud de las contracciones espontáneas.
El músculo liso de los esfínteres generalmente se contrae.
La retención de la orina se incrementa con las catecolaminas.
El músculo uterino en mujeres no preñadas se contrae en respuesta a la adrenalina, mientras que durante el embarazo se inhibe el tono y las contracciones de este músculo.
El músculo liso bronquiolar se relaja por acción de la adrenalina.
Respecto al músculo liso multiunitario, los músculos pilomotores se contraen por acción de la adrenalina, determinando la erección del pelo.
En el ojo, la adrenalina causa contracción del músculo radial del iris generando dilatación de la pupila, mientras que causa relajación del músculo ciliar produciendo un aplanamiento de las lentes, permitiendo que el ojo enfoque a objetos distantes.

Figura 9: Acciones de las catecolaminas sobre los ejes endocrinos.
Respecto a su acción en los ejes endocrinos (figura 9) se sabe que regulan el eje hipotálamo-hipófisis. No obstante las acciones sobre las hormonas implicadas en la respuesta al estrés son las siguientes:
– La disminución del flujo sanguíneo renal determina la liberación de renina por parte de las células yuxtaglomerulares. Esta renina determina el estímulo para la formación de angiotensina II y en consecuencia de aldosterona, evitándose así la pérdida de sodio, de líquido y la acumulación de potasio.
– En el páncreas endocrino decrece la liberación de insulina por parte de las células B, estimuladas por el incremento de glucosa en sangre; e incrementa la liberación de glucagón por parte de las células A.
– Estimula la liberación de PTH, aunque en situación de hipercalcemia no afecta a dicha liberación.
– En la glándula tiroides donde hay una abundante inervación simpática, se estimula la síntesis y liberación de hormonas tiroideas, causa por la cual también se puede explicar el efecto termogénico de las catecolaminas.
En los otros ejes hay evidencias de las acciones reguladoras de las catecolaminas aunque no se sabe aún su significado funcional.
Ante cualquiera de las situaciones fisiológicas que determinan una respuesta simpática, los eventos que se producen pueden resumirse de la siguiente manera:
1) Hiperglucemia
2) Incremento del Gasto Cardíaco
3) Incremento de la presión arterial media
4) Redistribución del flujo sanguíneo
5) Incremento de la temperatura por incremento del metabolismo en el tejido adiposo marrón e incremento en la liberación de hormonas tiroideas
6) Incremento del volumen líquido extracelular
7) Hiperlacticidemia
8) Disminución de la actividad motora gastrointestinal y genitourinaria
9) Incremento de la agudeza visual
10) Broncodilatación
11) Incremento en el consumo de oxígeno
La dopamina produce vasodilatación renal y en el mesenterio. Tiene efecto inotrópico positivo en el corazón (R-ß1). En dosis moderadas tiene un efecto neto de incremento de la presión sistólica sin cambios en la diastólica. También se ha visto que se sintetiza en la corteza renal y causa natriuresis.
REGULACIÓN DE LA SECRECIÓN DE LAS CATECOLAMINAS
La secreción medulo-adrenal es parte integral de la reacción provocada por la estimulación del sistema nervioso simpático. Así factores como la percepción, la anticipación a un daño (ansiedad), un trauma, el dolor, la hipovolemia por hemorragia o pérdida de líquido, la hipotensión, la anoxia, las temperaturas extremas, la hipoglucemia y el ejercicio intenso, entre otros, causan secreción medulo-adrenal.
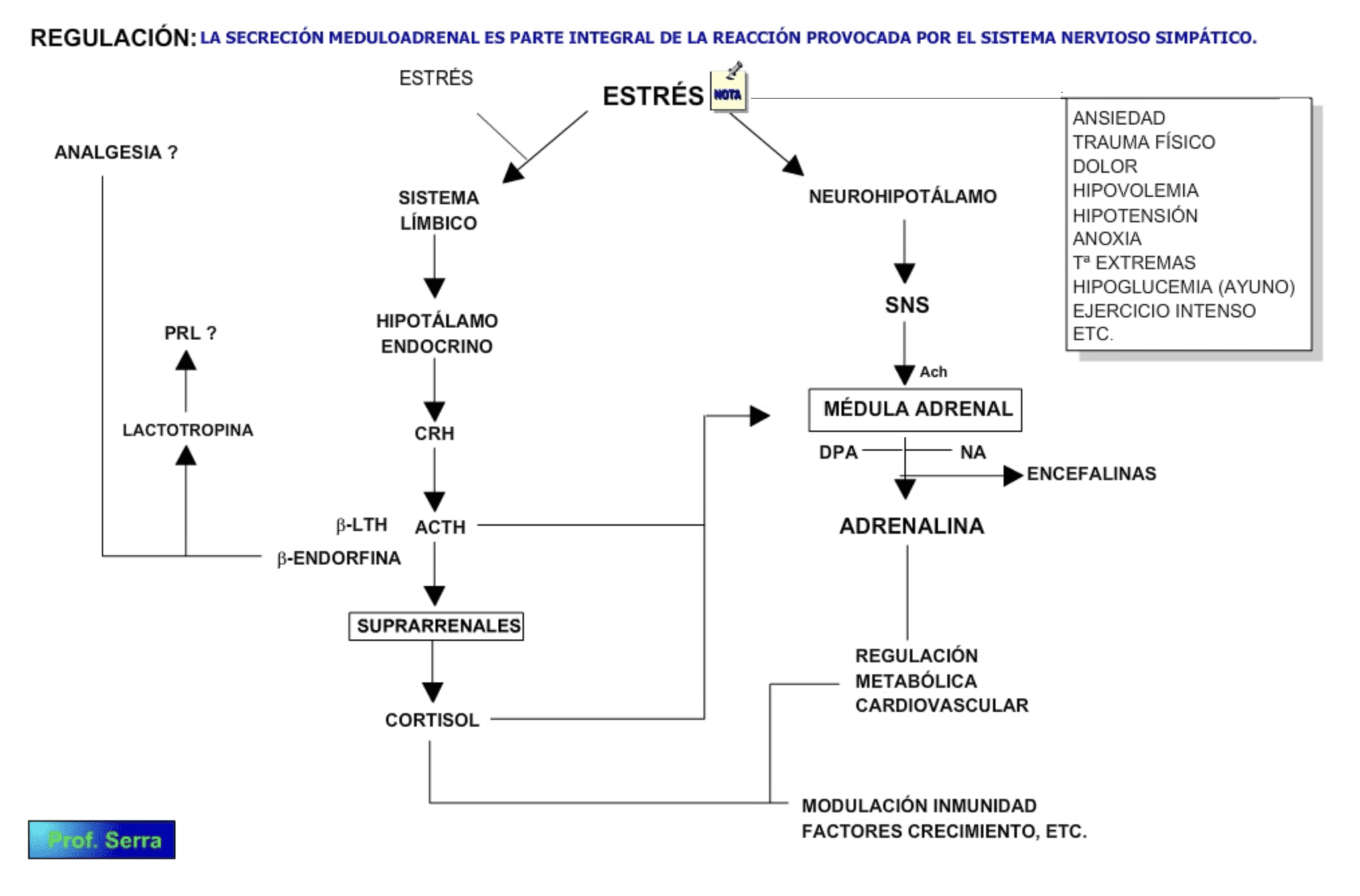
Figura 10: Regulación de la secreción de las catecolaminas.
La reacción simpática organizada en el hipotálamo llega a la médula a través de las terminaciones colinérgicas preganglionares. Esta acetilcolina despolariza a la célula cromafín induciendo la entrada de calcio, disparando la contracción de los microfilamentos y permitiendo por tanto, la liberación del contenido vesicular mediante exocitosis. Por otra parte, la ACTH estimula a la tirosín-hidroxilasa y a la dopamina hidroxilasa, activando así la síntesis de las catecolaminas. El cortisol también estimula a la metiltransferasa, favoreciendo por tanto la síntesis de adrenalina. En consecuencia la estimulación neuroendocrina de la glándula suprarrenal determina la liberación de todas sus hormonas, las cuales están implicadas en la reacción al estrés.
Al parecer la secreción de noradrenalina o adrenalina depende de la situación de estrés del sujeto. Así la primera se secreta en situaciones de estrés emocional familiares para el sujeto. Mientras que la adrenalina se secreta en situaciones donde el sujeto no sabe lo que viene.
En tumores cromafines, la histamina y el glucagón también estimulan la secreción de catecolaminas.
ALTERACIONES EN LA SECRECIÓN DE LAS CATECOLAMINAS
No se conocen síndromes de hiposecreción de estas hormonas en el adulto humano. En niños pequeños hay evidencias de una hipoglucemia idiopática por deficiencias de adrenalina, la cual se trata con preparados de larga acción hormonal.

Figura 11: Alteraciones en la secreción de las catecolaminas.
La hipersecreción (figura 11) generalmente se produce por tumores en las células cromafines (feocromocitomas). Las consecuencias más generales son: una hipertensión sostenida , en la mayoría de los pacientes, o esporádica en una minoría de estos pacientes. La aparición esporádica de esta hipertensión coincide con brotes intensos de secreción de adrenalina, provocados por un estrés, cambio postural, anestesia, etc., y conlleva dolor de cabeza intenso, palpitaciones, dolor torácico, ansiedad y sensación de muerte próxima, transpiración fría, palidez cutánea y visión borrosa, además de una hipertensión muy alta. La frecuencia cardiaca se verá aumentada si la hipersecreción es de adrenalina y disminuida si ésta es de noradrenalina.
Si la hipersecreción es mantenida, se pierde peso debido al incremento de la velocidad metabólica y la hiperglucemia. El tratamiento consiste en la extirpación del tumor medular y la aplicación de antagonistas alfa-adrenérgicos (fentolamina, prazosin, etc). El propranolol (antagonista ß-adrenérgico) es útil para reducir la taquicardia, pero su utilización requiere la administración simultánea de un antagonista alfa-adrenérgico, de lo contrario se incrementaría la hipertensión.
También aparece hipersecreción en los neuroblastomas (tumores de la cresta neural).
![]()
 Views Today : 751
Views Today : 751 Views This Month : 15897
Views This Month : 15897 Views This Year : 54352
Views This Year : 54352 Total views : 340415
Total views : 340415