Regulación del sistema respiratorio

Ingrese su ID de correo electrónico

CENTROS RESPIRATORIOS
Localización de los centro nerviosos implicados en la generación del ritmo respiratorio (Figura 2):
Mediante estudios de ablación y registros electrofisiológicos relacionados con la actividad respiratoria, se ha podido localizar la ubicación de los centros nerviosos implicados en la generación y regulación del ritmo respiratorio.

Figura-2: Centros reguladores de la respiración.
En el bulbo, en su formación reticular y por debajo del piso del cuarto ventrículo, se encuentran centros implicados directamente en la autogeneración del ritmo respiratorio básico y son:
1. El grupo respiratorio dorsal (GRD), implicado principalmente en la inspiración y que algunos consideran es responsable del ritmo respiratorio.
2. El grupo respiratorio ventral (GRV), relacionado principalmente en la espiración.
El primero (GRD) está localizado en los confines del núcleo del tracto solitario (NTS), con neuronas activas durante la inspiración. Con axones contralaterales que inervan a través del frénico el diafragma y axones contralaterales que van al GRV, inhibiendo a sus neuronas espiratorias. Éste recibe gran parte de la información sensorial respiratoria. Tiene actividad rítmica y es responsable del ritmo básico ventilatorio.
El segundo grupo (GRV) contiene dos poblaciones de neuronas unas en el núcleo ambiguo, con neuronas premotoras inspiratorias además de neuronas motoras de los músculos laríngeos y parasimpáticas a los bronquiolos y corazón. Otras en el retroambiguo, donde en su región rostral encontramos neuronas premotoras inspiratorias (I) con axones contralaterales hacia las Mns espinales que inervan a los músculos intercostales externos (inspiratorios) y músculos accesorios. En su región caudal, se encuentran neuronas premotoras espiratorias (E) que inervan de forma ipsilateral a las Mns espinales que inervan a los intercostales internos (espiratorios) y músculos abdominales. Las neuronas E parecen tener efectos inhibitorios durante la fase inspiratoria sobre las neuronas I, siendo así responsables de la finalización de dicha fase.
El complejo de Bötzinger (centro apnéusico) está compuesto exclusivamente por neuronas espiratorias que reciben entrada sensorial desde el NTS y tienen un efecto inhibitorio sobre las neuronas inspiratorias del GRD y GRV y un efecto excitatorio sobre las neuronas espiratorias del GRV.
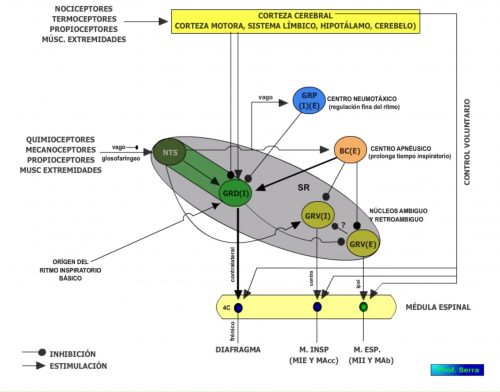
Figura-3: Organización neuronal de los centros reguladores respiratorios.
La información sensorial del vago es de tipo inhibidor sobre las neuronas inspiratorias.
En el puente tenemos:
Neuronas del grupo respiratorio del puente (GRP) (antiguamente conocidas como centro neumotáxico) son de tipo inspiratorio localizadas en el núcleo parabraquial lateral (NPL) y núcleo de Kölikker-Fuse (NKF) y espiratorio en el núcleo parabraquial media (NPM). Parece implicado en la regulación de la profundidad inspiratoria y recibe información sensorial vagal. Los dos primeros regulan el ritmo inspiratorio actuando sobre el GRD.
ESTOS GRUPOS ORGANIZAN LA INFORMACIÓN NECESARIA PARA GENERAR EL RITMO RESPIRATORIO Y LA PROFUNDIDAD DE LA INSPIRACIÓN (Figura 3), SIENDO EL GRUPO RESPIRATORIO VENTRAL DONDE SE ORIGINA EL RITMO Y EL RESTO SE ENCARGAN DE MODULARLO.
REFLEJOS RESPIRATORIOS
EL CONTROL del ritmo y profundidad respiratoria es la consecuencia de la integración de todas las aferencias que llegan a los centros respiratorios bulbares, donde se origina el ritmo básico respiratorio.

Figura-4: Organización de los reflejos pulmonares.
El objetivo de este control radica en encontrar la combinación óptima de frecuencia y volumen ventilatorio para lograr la ventilación con esfuerzo mínimo. Y es muy importante el estado de vigilia del sujeto.
AFERENCIAS SENSORIALES EN EL CONTROL DEL APARATO RESPIRATORIO:
La información sensorial que llega a los centros respiratorios para modular la frecuencia y profundidad respiratoria se pueden clasificar en tres, de acuerdo con el tipo de información que llevan:
1. Información química: procedente de los quimioceptores periféricos ubicados en la circulación arterial, donde se miden los niveles plasmáticos de O2, CO2 y el pH; y los centrales, ubicados en el suelo del cuarto ventrículo y miden el CO2 del líquido cefalorraquídeo.
2. Información propioceptora: que traen información de la actividad muscular tanto de los músculos respiratorios, como de los músculos de las extremidades.
3. Información mecanoceptora: procedente de los barorreceptores ubicados en la región pulmonar y también en el sistema cardiovascular.
Otras informaciones sensoriales también ejercen su efecto respiratorio pero no de forma directa, sino a través de centros superiores ubicados en el hipotálamo, sistema límbico y córtex cerebral, donde se coordinan respuestas integradas que afectan a la función del sistema respiratorio.


SENSORES QUÍMICOS PERIFÉRICOS
Cuerpos carotídeos y cuerpos aórticos.
Los cuerpos carotídeos se encuentran situados en la bifurcación de las carótidas, con inervación sensorial del glosofaríngeo (IX) y un flujo sanguíneo de 2000 ml/100 g tejido/min. (33 veces más que el cerebro).
Miden el O2 disuelto. (Figura 5)
Histológicamente son células sensoriales tipo I o células glómicas rodeadas por células gliales (tipo II).
El neurotransmisor es la dopamina.
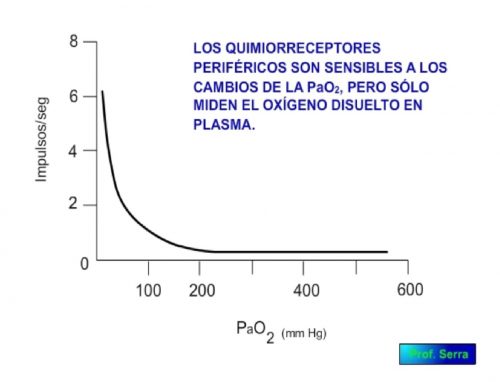
Figura-7: Cambios de ventilación frente al oxígeno disuelto.
El principal estímulo es la PaO2 seguido de la PaCO2 y el pH (Figura 6), aunque estos últimos requieren cambios importantes para ejercer un efecto.
Como se puede apreciar en la figura 7, estos sensores trabajan en el rango fisiológico de PaO2 de forma que por encima de una PaO2 de 200 ya no disparan. Y por debajo de una PaO2 de 40, la frecuencia de disparo es máxima.
SENSORES QUÍMICOS CENTRALES
En la base del cuarto ventrículo cerebral (en el bulbo) se encuentran neuronas sensibles a los cambios del CO2 en el líquido cefalorraquídeo (LCR) (Figura 8). Estas neuronas están en estrecha relación con los "centros respiratorios". Realmente son sensibles a los cambios en la concentración de hidrogeniones provocados por los cambios en la presión de CO2, el cual pasa fácilmente la barrera hematoencefálica.

Figura-17: Cambios de la ventilación por cambios en el CO2 cefalorraquídeo.
El CO2 una vez en el LCR se hidrata y forma CO3H– y H+ este incremento en hidrogeniones es el causante del incremento en la actividad de las neuronas inspiratorias. La respuesta es muy rápida, menor de 1 minuto, pero tiene un límite, de forma que a valores de PCO2 mayores de 70 mm Hg se produce un efecto paralizante de los centros respiratorios.
Si se hiperventila durante un tiempo, se puede mantener más tiempo suspendida la respiración.
INTERACCIONES ENTRE ELLOS
Cuando se utilizan drogas que disminuyen la sensibilidad del sensor central al CO2 (barbitúricos, morfina, etc.) se pone de manifiesto la regulación mediada por los quimioceptores periféricos. (Figura 9)
Si analizamos el efecto conjunto del O2 y CO2 en la regulación química del aparato respiratorio obtenemos los gráficos de la figura, donde podemos observar lo siguiente:
A medida que aumenta el valor de la PaCO2 se produce un incremento de la ventilación, que se hace máximo cuando los valores de la PaO2 caen por debajo de los valores vitales.

Figura-9: Interacción entre los sensores químicos.
Cuando analizamos los cambios de ventilación con respecto a la PaO2 observamos que son poco importantes y muy dependientes de los valores de la PaCO2, de forma que si estos últimos son mayores de 40 mm Hg se produce un aumento de la ventilación a medida que baja la PaO2, pero si están por debajo de 40, se necesita una caída importante de la PaO2 para que se produzca un aumento importante de la ventilación [1]El hecho de que entre los valores de 60 a 100 mm Hg en la PA O2 no se produzca un cambio importante en la ventilación se puede explicar por la acción cooperativa de dos efectos: la disminución de la PCO2 como consecuencia del ligero aumento de la ventilación, lo que supone inhibición de la misma; y el aumento de la desoxiHb que por ser un ácido débil hace subir el pH sanguíneo y consiguiente depresión de la ventilación.El hecho … Continúe leyendo.
La detención voluntaria de la respiración lleva a un incremento de la PaCO2 y caída de la PaO2 hasta un punto donde se produce un estímulo de los centros respiratorios por encima del control voluntario: Punto de ruptura. Este punto se puede atrasar si se seccionan los cuerpos carotídeos. También se puede alargar con hiperventilaciones previas, que bajen el valor de la PaCO2.
Consecuencia de este análisis es que el parámetro químico que gobierna la ventilación es la PaCO2 y que sólo se hace visible el papel de la PaO2 cuando éste alcanza valores que ponen en peligro la supervivencia celular.
En los trastornos del equilibrio ácido-base se observan cambios respiratorios.
En la acidosis metabólica (provocada por acumulación de ácidos no volátiles, por ejemplo cuerpos cetónicos (diabetes sacarina)) se produce un incremento de la ventilación (respuesta de Kussmaul).
En la alcalosis metabólica (por ejemplo, vómitos prolongados con pérdida de ClH) se produce disminución de la ventilación.
Por otra parte las modificaciones ventilatorias independientes a los cambios del equilibrio ácido-base ejercen cambios en éste:
La hiperventilación produce alcalosis respiratoria.
La hipoventilación produce acidosis respiratoria.







![]()
 Views Today : 207
Views Today : 207 Views This Month : 1838
Views This Month : 1838 Views This Year : 27461
Views This Year : 27461 Total views : 313524
Total views : 313524