

Ingrese su ID de correo electrónico
LA SS Y EL EJE SOMATOTRÓPICO
El eje somatotrópico comienza, como muchos otros ejes, en el hipotálamo endocrino. Ahí, neuronas del núcleo arqueado y del hipotálamo anterior, envían axones somatotrópicos que liberan SST a los capilares de la eminencia media y de ahí a las células somatotrófas de la adenohipófisis, Ésta SST ejerce un efecto inhibitorio sobre la secreción de la GH.
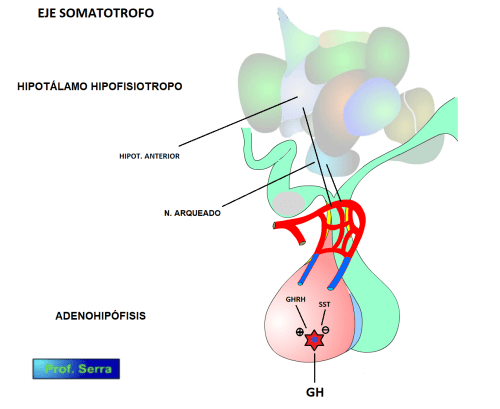
Figura 1: Eje somatotrofo.
LA SST. ESTRUCTURA, LOCALIZACIÓN Y PRODUCCIÓN
La somatostatina es una hormona peptídica que procede de una preprohormona de 116 aa (cromosoma 3 188.87 – 188.87 Mb ). Por clivación da dos GHIH, uno de 14 aa (SST-14) y otro de 28 aa (SST-28). En 1999 se identificó una variante de la somatostatina, denominada costistatina-14 por su permanencia en el córtex cerebral, pero también en células del sistema inmune como monocitos, macrófagos y células dendríticas (Weinstock & Elliott 2000 [1]Weinstock JV& Elliott D 2000 The somatostatin immunoregulatory circuit present at sites of chronic inflammation. European Journal of Endocrinology 143(Suppl 1) S15–S19.[Texto libre]). Se une a los mismos receptores que la SS-14 y también al receptor de la ghrelina (Broglio et al. 2002 [2]Broglio F, Arvat E, Benso A, Gottero C, Prodam F, Grottoli S, Papotti M, Muccioli G, van der Lely AJ, Deghenghi R et al. 2002 Endocrine activities of cortistatin-14 and its interaction with GHRH and ghrelin in humans. Journal of Clinical Endocrinology and Metabolism 87 3783–3790.[Texto completo].
La hormona somatostatina presenta dos formas activas, una con 28 y otra de 14 aminoácidos. La distribución, abundancia, o presencia de éstas, dependerá del tejido donde se encuentre. En el tejido pancreático o el hipotálamo presentan única, o principalmente, formas de 14 aminoácidos. En el tejido intestinal, por su parte, su composición es mayormente de 28 aminoácidos.

Figura 2. Estructuras químicas de las somatostatinas 14 y 28.
El descubrimiento de esta hormona fue realizado por Krulich y colaboradores en el año 1968 [3]Krulich L., Dhariwall A.P.S. y McCann S.M. Stimulatory and inhibitory effects of purified hypothalamic extracts on growth hormone release from rat pituitary in vitro. Endocrinol (1968) 83:783-790. en el hipotálamo ovino. Mientras investigaban el factor que liberaba la hormona del crecimiento, estos investigadores determinaron que extractos del hipotálamo inhibían la secreción hipofisaria.
Posteriormente, Brazeau y colaboradores, en el año 1973 [4]Brazeau P, Vale W, Burgus R, Ling N, Butcher M, Rivier J, Guillemin R. Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 1973; 179:77-79., la caracterizaron en el laboratorio de Roger Guillemin, donde intentaron aislar, a partir de células de hipófisis, el factor responsable de la liberación de la hormona del crecimiento (GH); en su lugar, aislaron un tetradecapéptido que inhibía la secreción de dicha hormona tanto in vivo como in vitro. Por este descubrimiento, Guillemin fue galardonado con el Premio Nobel de Medicina y Fisiología en 1977. Al tetradecapéptido aislado se le bautizó finalmente como “factor inhibidor de la liberación de la somatotropina” (SRIF, por sus siglas en inglés) o somatostatina (SST), término más usado actualmente, aunque no englobe sus múltiples acciones. Estudios posteriores demostraron que la inhibición de la liberación de la somatotropina es sólo uno de los múltiples efectos de la SST. Las funciones de la SST se centran en modular la neurotransmisión, secreción glandular, contractibilidad de la musculatura lisa y proliferación celular.
SchalIy y cols. [5]SCHALLY, A. V.; A.; ARIMURA, A.; REDDING, E. W.; Linthicum, G. L.: Isolation 01′ porcine GHcreleasing inhibiting hormane. The existence of trhee formas of GHRIH. Federation Proceedings 34:586; 1975. en 1975 aislaron un péptido extraído de hipotálamos porcinos con capacidad de inhibir la liberación de hormona de crecimiento.
En el año 1977 Pelletier y colaboradores [6]PELLETIER, G.; LABRIE, F.; ARIMURA, A.; AND SCHALLY, A. V.: Electron microscopic inmuno histochemical localization of growth hormone release inhibitng hormone (somatostatin) in the rat median eminenee. American Joumal of Anatomy 140: 445; 1974. descubrieron en el hipotálamo humano la somatostatina (SST).
A partir de entonces esta hormona ha sido observada en casi todos los sistemas de tejidos y órganos. Está presente no solo en humanos sino también en otros mamíferos y vertebrados.
Se identifica, además de en el hipotálamo hipofisario, en el córtex, en el sistema nervioso periférico, intestino (plexo mientérico y submucoso) y páncreas exocrino (células delta).
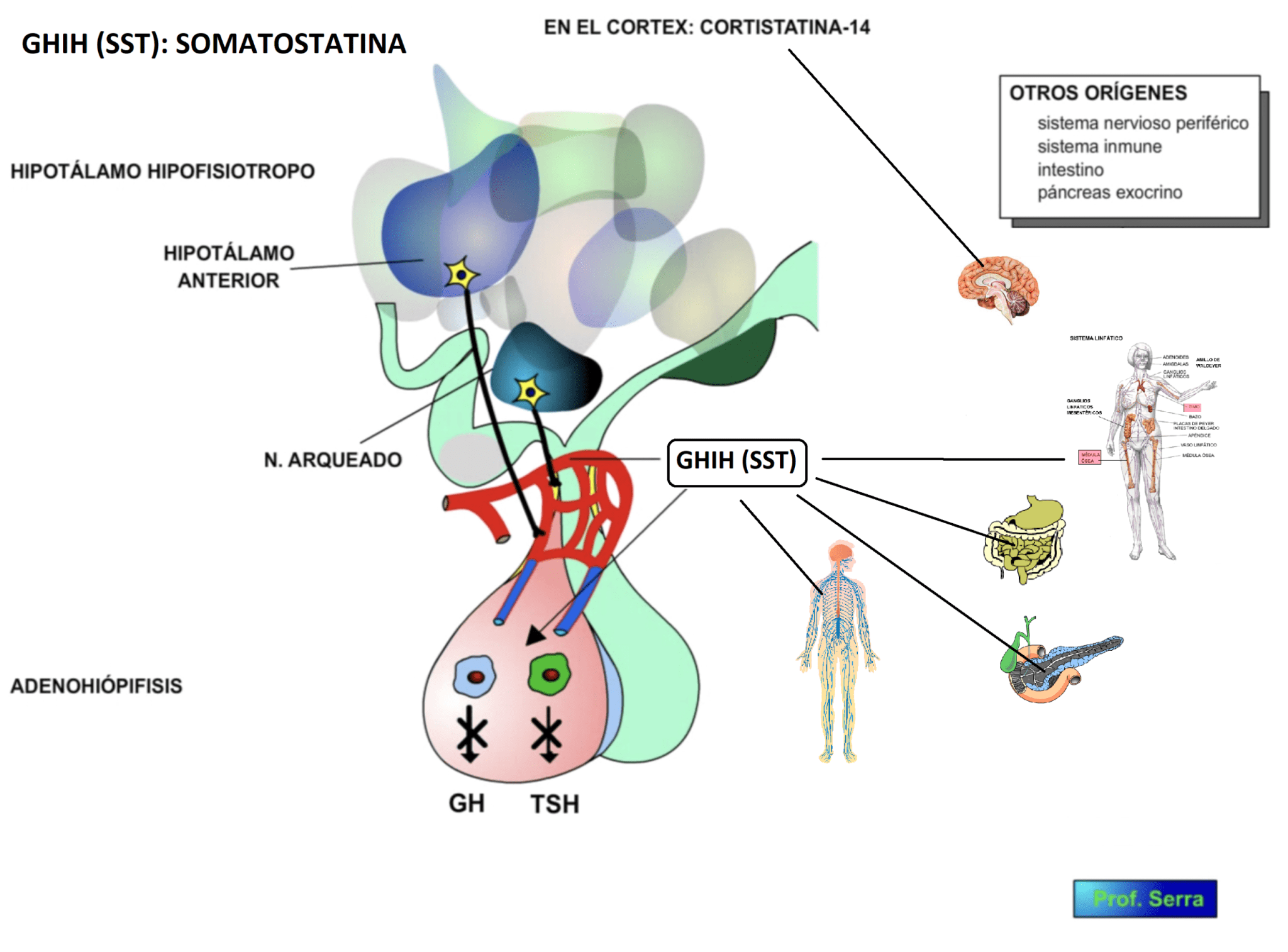
Figura 3: Orígenes de la SST
Respecto a su transcripción genética está regulada por una gran variedad de moléculas, como la glucosa, hormonas tiroideas, serotonina, péptido intestinal vasoactivo, factor de crecimiento similar a la insulina 1 IGF-1, etc. No se conoce el mecanismo molecular mediante el cual actúan pero sí que la activación de la transcripción está directamente mediada por el AMPc, vía CRE. Después de la estimulación de la enzima adenilato ciclasa (AC), inducida por vía receptor, se produce un aumento en los niveles de AMPc, que activan a una proteína quinasa (PKA), cuya subunidad catalítica de la será transportada al núcleo celular donde, junto al coactivador CBP (proteína de unión a CREB) se fosforila una proteína nuclear denominada CREB (proteína de unión a CRE). La proteína CREB fosforilada se une al elemento CRE del gen de la SST, estimulando su transcripción.
LA SST. RECEPTORES
Rn-GRH (Gi/AMPc). Cinco tipos de receptores.(CorletoV. D. et al. 2004)[7]CorletoV. D., S. Nasoni, F. Panzuto, S. Cassetta and G. Delle Fave. Somatostatin receptor subtypes: basic pharmacology and tissue distribution. Digestive and Liver Disease Volume 36, Supplement 1, February 2004, Pages S8-S16 [Texto libre]) con isoformas. Codificados como: SSTR1, SSTR2, SSTR3, SSTR4 y SSTR5 [8]Hofland, L. and Lamberts, S. (1996). Somatostatin receptors and disease: Role of receptor subtypes. Baillière’s Clinical Endocrinology and Metabolism, 10(1), pp.163-176..
Esta diversidad de receptores explica la multiplicidad de acciones biológicas que se han observado para la somatostatina. Un mismo subtipo de receptor puede inducir respuestas biológicas distintas o bien una respuesta biológica puede implicar más de un subtipo de receptor. Los subtipos de receptores 1 al 4 (SSTR1-4) se unen con alta afinidad tanto a la somatostatina 14 como a la somatostatina 28; sin embargo, el subtipo de receptor 5 (SSTR5) tiene mayor afinidad por la somatostatina 28.
Estos receptores se han identificado en un gran número de órganos: cerebro, hipófisis, aparato digestivo, placenta, páncreas endocrino y exocrino, adrenales, tiroides, riñones y células inmunes. En la glándula pituitaria de un sujeto sano se encuentran los siguientes receptores: SSTR1, 2 y 5 y probablemente el 3 [9]Ben-Shlomo, A., Liu, N. and Melmed, S. (2016). Somatostatin and dopamine receptor regulation of pituitary somatotroph adenomas. Pituitary.. El receptor SSTR2 es el principal mediador de la inhibición de la secreción de la GH.
El mecanismo de acción de estos cinco subtipos e isoformas está ligado a diferentes vías de transducción, entre ellas: adenilato ciclasa, fosfolipasa C, fosfolipasa A2, guanilato ciclasa, canales iónicos y tirosina fosfatasa, fosfotirosina fosfatasas (PTPs), proteínas quinasas activadas por mitógenos (MAPK) y la bomba Na+ /H+ [10]Patel, Y. (1999). Somatostatin and Its Receptor Family. Frontiers in Neuroendocrinology, 20(3), pp.157-198..
Los receptores de somatostatina, pertenecen a la superfamilia de los GPCR (receptor acoplado a proteína G). Poseen siete dominios hidrofóbicos α-helicoidales transmembrana, de 20 a 25 aminoácidos de longitud, separados por bucles extra- e intracelulares de residuos hidrofílicos, con una secuencia N-terminal extracelular y una secuencia C-terminal intracelular. Los genes que codifican los cinco subtipos de receptores de somatostatina están localizados en distintos cromosomas. Se ha demostrado que una vez producida la activación de los receptores por la unión de su agonista, éstos se desensibilizan mediante la fosforilación de residuos de serina y treonina presentes en el dominio C-terminal intracelular del receptor, induciendo el desacoplamiento entre el receptor y las proteínas G. Esta fosforilación se lleva a cabo por miembros de la familia de quinasas de GPCRs (GRKs). La fosforilación del receptor y el posterior reclutamiento de proteínas citosólicas denominadas arrestinas producen el desacoplamiento entre el receptor y las proteínas G, y la consiguiente internación y posterior degradación del receptor. Todos los receptores, excepto el SSTR4, se desensibilizan mediante el mecanismo descrito anteriormente.
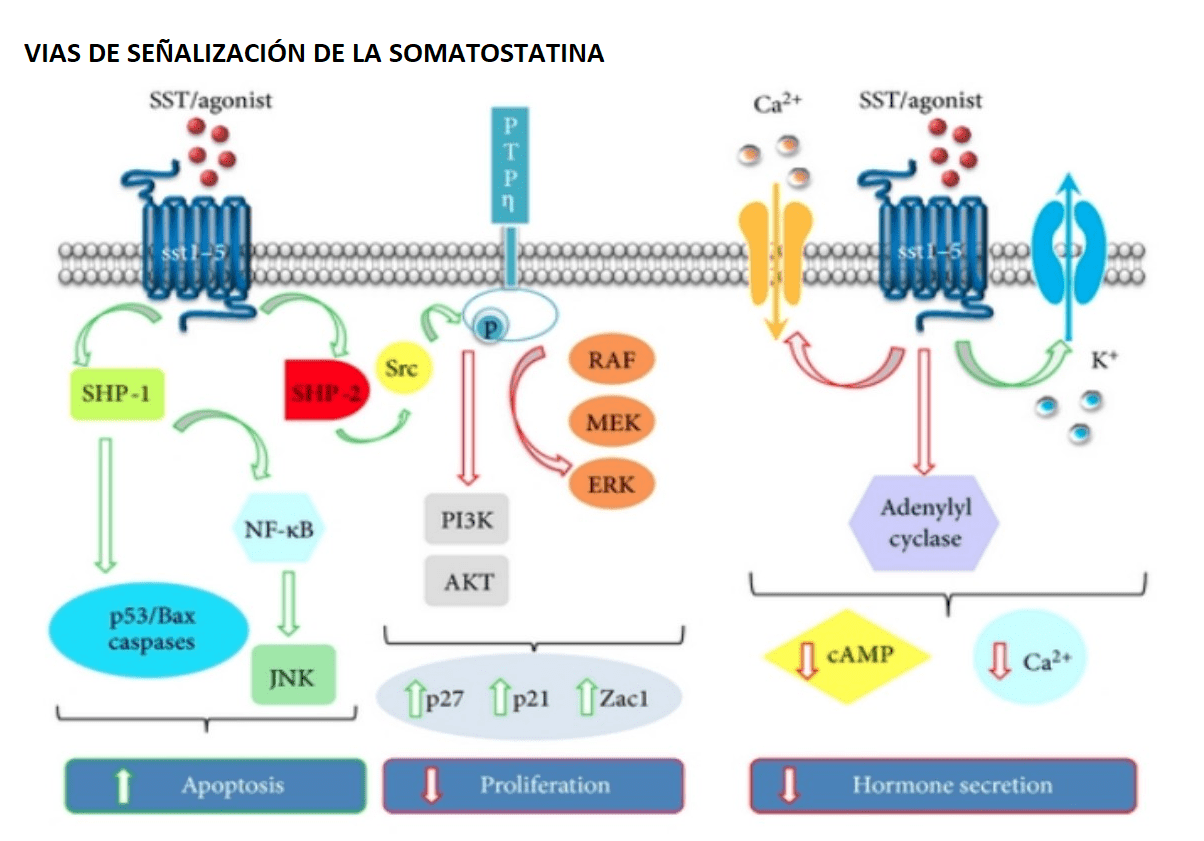
Los efectos que la SST ejerce en sus células diana son básicamente dos:
- Inhibición de la secreción. Mediante:
- Inhibición de la enzima adenilato ciclasa (AC).
- Estimulación de los canales de K+ dependientes de voltaje.
- Inhibición de los canales de Ca2+ dependientes de voltaje.
- Inhibición de la exocitosis, mediado por el acoplamiento de los receptores de somatostatina a vesículas de secreción.
- Inhibición de la proliferación celular. Mediante:
- Inhibición de la bomba Na+ /H+
Estimulación de una fosfotirosina fosfatasa (PTP) de membrana.
Figura 4. Vías de señalización de la SST en sus tejidos diana. [11]La adenilato ciclasa es una enzima de la membrana plasmática que convierte el ATP en AMPc, un segundo mensajero que regula numerosas de funciones biológicas en la célula. La actividad de esta enzima está controlada por receptores estimuladores e inhibidores, a través de proteínas Gs y Gi, respectivamente. El principal papel de las proteínas G es acoplar los receptores a los múltiples efectores regulados por la somatostatina. Estas proteínas G son … Continúe leyendo
LA SST. FUNCIONES
La somatostatina puede actuar de las siguientes formas:
1. Neurohormona: la somatostatina es liberada desde el hipotálamo-hipofisario al sistema portahipofisario e inhibe la secreción de hormonas pulsátiles anterohipofisarias.
2. Hormona periférica: la somatostatina también es secretada por el aparato
digestivo e inhibe las secreciones hormonales pancreáticas (insulina, glucagón).
3. Neurotransmisor: la somatostatina ejerce una acción inhibitoria de la actividad de otras neuronas del sistema central y autónomo.
4. Factor Paracrino: factor de regulación local de actividades secretorias a nivel del páncreas o de la proliferación celular. Ejerce sus actividades en numerosos puntos del tubo digestivo.

Figura 5. Funciones de la somatostatina.
En el eje hipotálamo- hipofisario es capaz de actuar sobre la liberación de la GH y tirotropina (TSH).[12]MOREL, G., MESGUICH, P., DUBOIS, M.P. and DUBOIS, M.P. :Ultrastructural evidence for endogenous somatostatin-like immunoreactivity in the pituitary gland. Neuroendocrinology 36: 291-299, 1983. .
HORMONA DEL CRECIMIENTO (GH)
El primer efecto biológico conocido de la somatostatina fue la inhibición de la secreción de GH en cultivos de células hipofisarias (BRAZEAU y cols., 1973)[13]BRAZEAU, P., VALE, W . , BURGUS, R., LING, N., BUTCHER, M. , RIVIER, J. and GUILLEMIN, R .: Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179: 77-79, 1973.. La somatostatina, efectivamente, es capaz de suprimir la respuesta de la GH al ejercicio (PRANGE-HANSEN y cols., 1973)[14]PRANGE-HANSEN, A., ORSKOV, H., SEYER-HANSEN, K. and LUNDBAEK, K .: Some actions of growth hormonerelease inhibiting factor. Br. Med. J. 3: 522-524, 1973., a la hipoglucemia insulínica (HALL y cols., 1973)[15]HALL, R., BESSER, G.M., SCHALLY, A.V., COY, D.H., EVERED, D. , GOLDIE, D.J., KASTIN, A.J., McNEILLY, A. S., MORTIMER, C.H., PHENEKOS, C., TURNBRIDGE, W.M.G. and WEIGHTMAN, D.: Actions of growth hormone-release inhibiting hormone in healthy men and acromegaly. Lancet 2: 581-584, 1973., a la L-dopa (SILER y cols., 1973)[16]SILER, T.M., VANDERBERG, G. , YEN, S.S.C., BRAZEAU, P., VALE, W. and GUILLEMIN, R . : Inhibition of growth hormone release in humans by somatostatin. J. Clin. Endocrinol. Metab. 37; 632-634, 1973. , arginina (MORTIMER y cols., 1974)[17]MORTIMER, C.H., TUNBRIDGE, W.M.G., CARR, D., YEOMANS, L. , LIND, T., COY, D.H., BLOOM, S.R., KASTIN, A., MALLINSON, C.N., BESSER, G.M., SCHALLY, A.V. and HALL, R .: Effects of growth hormonerelease inhibiting hormone on circulating glucagon, insulin and growth hormone in normal, diabetic, acromegalic and hypopituitary patients. Lancet 1: 697-701, 1974., estimulación eléctrica de los núcleos amigdaloides ventromedial y basolateral del hipotálamo (MARTIN, 1974)[18]MARTIN, J.B.: Inhibition effect of somatostatin (SRIF) on the release of growth hormone induced in the rat by electrical stimulation. Endocrinology 94: 497-502, 1974. , morfina, neurotensina y sustancia P (BROWN y VALE; 1975)[19]BROWN, M. and VALE, W .: Growth hormone release in the rat : effects of somatostatin and thyrotropinreleasing factor. Endocrinology 97: 1151-1156, 1975. , dibutiril-AMPc (PERACCHI y cols., 1976)[20]PERACCHI, M., RESCHINI, E., CANTALAMESSA, L, CATANIA, A. and GUISTINA, G. : Inhibitory effect of somatostatin on dibutyryl cyclic AMP-induced insulin and growth hormone release in human subjects. Metabolism 25; 321-328, 1976. , succión del pezón (CHEN y cols., 1974)[21]CHEN, H.J., MUELLER, G.P. and MEITES, J .: Effects of L-dopa and somatostatin on suckling-induced release of prolactin and growth hormone. Endocrine Res. Commun. 1: 283-291, 1974. , alimentos (BESSER y cols., 1974b)[22]BESSER, G.M., MORTIMER, C.H., McNEILLY, A,S., THORNER, M. , BATISTONI, G., BLOOM, S., KASTRUP, K. , HANSSEN, K., HALL, R., COY, D., KASTIN, A. and SCHALLY, A.: Long term infusion of growth hormone-release inhibiting hormone in acromegaly: effects on pituitary and pancreatic hormones. Br. Med. J. 4 : 622-627, 1974b. y prostaglandinas E2 (GIULIANO y cols., 1979[23]GIULIANO, D., TORELLA, R., SGAMBATO, S. and D’ONOFRIO, E.: Effects of alfa- and beta-adrenergic inhibition and somatostatin on plasma glucose, free fatty acids, insulin, glucagon and growth hormone responses to prostaglandin E\ in man. J. Clin. Endocrinol. Metab. 48: 302-308, 1979. ; SZABO y FROHMAN, 1975[24]SZABO, M . and FROHMAN, L.A.: Effects of porcine stalk median eminence and prostaglandin E2 on rat growth hormone secretion in vivo and their inhibition by somatostatin. Endocrinology 96: 955-961, 1975. ). La somatostatina también inhibe los picos episódicos de GH que se observan durante el sueño (PARKER y cols., 1974[25]PARKER, D.C., ROSSMAN, L.G., SILER, T.M., RIVIER, J., YEN, S. and GUILLEMIN, R.: Inhibition of the sleep-related peak in physiologic human growth hormone release by somatostatin. J. Clin. Endocrinol. Metab. 38: 496-499, 1974. ; LUCKE y cols., 1976[26]LUCKE, C. , HOFFKEN, B. and VON ZUR MUHLEN, A. : Studies on the postponed growth hormone secretion following the infusion of somatostatin. Acta Endocrinol. 82: 460-466, 1976. ) y la vigilia (WARD y cols., 1975[27]WARD, F.R., LEBLANC, H. and YEN, S.S.C.: The inhibitory effect of somatostatin on growth hormone, insulin and glucagon secretion in diabetes mellitus. J. Clin. Endocrinol. Metab. 41: 527-632, 19751). Mas recientemente se ha podido demostrar que la somatostatina impide el efecto estimulador del GRF (GHRH) sobre las células somatotrópicas de la hipófisis (WEHRENBERG y cols., 1982[28]WEHRENBERG, W.B., LING, N. , BOHLEN, P., ESCH, F. , BRAZEAU, P. and GUILLEMIN, R .: Physiological roles of somatocrinin and somatostatin in the regulation of growth hormone secretion. Biochem. Biophys. Res. Commun. 109: 562-567, 1982. ).
TIROTROPINA.
En el eje tiroideo actúa inhibiendo la liberación de tirotropina (TSH) en preparaciones hipofisarias aisladas (VALE y cols., 1975[29]VALE, W. , BRAZEAU, P., RIVIER, C., BROWN, M. , BOSS, B. , RIVIER, J., BURGUS, R. , LING, N. and GUILLEMIN, R .: Somatostatin. Rec. Prog. Horm. Res. 31; 365-397, 1975.) y reduce los niveles basales de esta hormona en la rata (CHIHARA y cols. , 1978[30]CHIHARA, K., ARIMURA, A., CHIHARA, M. and SCHALLY, A.V.: Studies on the mechanism of growth hormone and thyrotropin response to somatostatin antiserum in anesthetized rats. Endocrinology 103; 1916- 1923, 1978.) y en el hombre (WEEKE y cols., 1975[31]WEEKE, J., HANSEN, A.P. and LUNDAEK, K.: Inhibition by somatostatin of basal levels of serum thyrotropin in normal men. J. Clin. Endocrinol. Metab. 41: 168-171, 1975. ).
La somatostatina es capaz también de inhibir la respuesta de la TSH a la TRH de una manera no competitiva (HALL y cols., 1973[32]HALL, R., BESSER, G.M., SCHALLY, A.V., COY, D.H., EVERED, D. , GOLDIE, D.J., KASTIN, A.J., McNEILLY, A. S., MORTIMER, C.H., PHENEKOS, C., TURNBRIDGE, W.M.G. and WEIGHTMAN, D.: Actions of growth hormone-release inhibiting hormone in healthy men and acromegaly. Lancet 2: 581-584, 1973.). Investigaciones ulteriores mostraron un efecto inhibidor de la somatostatina in vitro sobre la respuesta de la TSH a la teofilina (VALE y cols., 1974[33]VALE, W., RIVIER, C., BRAZEAU, P. and GUILLEMIN, R.: Effect of somatostatin on the secretion of thyrotropin and prolactin. Endocrinology 95: 968-977, 1974. ), monobutiri1-AMPc y prostaglandina E (BELANGER y cols., 1974[34]BELANGER, A., LABRIE, F., BORGEAT, P., SAVARY, M. , COTE, J. and BROUIN, J.: Inhibition of growth hormone and thyrotropin release by growth hormone release inhibiting hormone. Molec . Cel. Endocrinol. 1: 329-339, 1974.).
TRACTO DIGESTIVO
La SST inhibe la liberación de hormonas como la gastrina, secretina, colecistoquinina (CCK), ghrelina, motilina, péptido intestinal vasoactivo (VIP), péptido inhibidor gástrico (GIP), neurotensina y pepsina (SCHUSDZIARRA y cols., 1978)[35]SCHUSDZIARRA, V., IPP, E. , HARRIS, V,, DOBBS, R.E, RASKIN, P., ORCI, L. and UNGER, R.M.: Studies of physiology and pathophysiology of pancreaticD cell. Metabolism 27: 1227-1232, 1978. .
Al nivel pancreático, actúa como un potente inhibidor de la liberación de hormonas como la insulina, el glucagón o el polipéptido pancreático, participando de este modo en la regulación de la homeostasis metabólica.
La somatostatina también puede retrasar el vaciado gástrico tardío, acelerar el vaciado gástrico temprano y aumentar el tiempo de tránsito por el intestino delgado. Inhibe la motilidad de la vesícula biliar.
En glándulas exocrinas como las glándulas salivales y mucosa intestinal, la SST ejerce su papel inhibidor (ADLER y cols., 1980 [36]ADLER, G. , KOCH, A. and KERN, H.F.: Effect of somatostatin on rat exocrine pancreatic secretory process in normal and diseased state. Z. Gastroenterol. 418-426, 1980.).
Respecto al flujo sanguíneo mesentérico, la SST lo disminuye e incrementa la resistencia vascular mesentérica (REICHLIN, 1983[37]REICHLIN, S.: Somatostatin. N. Engl. J . Med. 309: 1495-1501, 1556-1563, 1983. ).
Disminuye la absorción intestinal de calcio, glucosa, galactosa, xilol, fructosa, lactosa, aminoácidos, triglicéridos y agua.
SISTEMA NERVIOSO CENTRAL:
La SST inhibe la liberación de dopamina y norepinefrina, causando así efectos sobre funciones sensoriales, locomotoras y cognitivas. De hecho, cambios en los niveles de somatostatina y sus receptores a este nivel están relacionados con enfermedades neurodegenerativas, tales como el Alzheimer o el Parkinson.
La aplicación de somatostatina disminuye la excitabilidad espontánea y evocada de un cierto número de neuronas de las cortezas cerebral y cerebelosa, hipotálamo y médula espinal (RENAUD, 1979[38]RENAUD, L.P.: Electrophysiology of brain peptides. En; GOTTO, A.M., Jr., PECK, E.J., Jr. and BOYD, A.E. , III (eds.): Brain peptides: a new endocrinology. Elsevier/North Hoiland, Amsterdam, Elsevier/North Holland, 1979; pp. 119-138. ). Esta acción, que en algunos aspectos recuerda a la del neurotransmisor inhibidor ácido gamma-aminobutírico (GABA), no era bloqueada por los antagonistas del GABA.
La somatostatina, además, puede antagonizar las convulsiones inducidas por la estricnina e incrementar la duración de la anestesia barbitúrica (McCANN, 1982[39]McCANN, S.M.: Physiology and somatostatin. Ann. pharmacology of Rev. Pharmacol. LHRH and Toxicol. 22: 491-515, 1982.). Sin embargo, otros autores han encontrado que la somatostatina ejerce una acción excitadora sobre la actividad eléctrica espontánea de las neuronas (DICHTER y DELFS, 1981[40]DICHTER, M.A. and DELFS, J.R.: Somatostatin and cortical neurons in cell culture. Adv. Biochem. Psychopharmacol. 28: 145-157, 1981. ), tal vez mediada a través de una inhibición de neuronas inhibidoras que provocaría la descarga de otras neuronas. Algunos aspectos de la sensibilidad se ven también afectados por la somatostatina. En las raíces de los ganglios dorsales el péptido parece modificar los impulsos dolorosos dirigidos centralmente mediante el control de la liberación de sustancia P en la zona de entrada de la raíz dorsal de la médula espinal (REICHLIN, 1981a[41]REICHLIN, S.: Neuroendocrinology. En: WILLIAMS, R.H. (ed.): Textbook of Endocrinology, 6th ed. Saunders, Philadelphia, 1981a; pp. 589-645.).
SISTEMA INMUNE: también se encuentra regulado por la SST, controlando negativamente la respuesta secretora de basófilos, monocitos e inmunoglobulinas y regulando la proliferación de linfocitos T y B, de un modo dosis-dependiente. Este tipo de regulación, también se ha observado en otros tejidos como el cardiovascular, donde la alta o baja concentración de somatostatina provoca una respuesta positiva o negativa de la contracción ventricular de los cardiomiocitos.
También tiene relación con procesos de regulación tumoral o en enfermedades de gran incidencia en la población.
OTRAS ACCIONES.
Inhibe la liberación de calcitonina y de hormona paratiroidea (HARGIS y cols., 1978 [42]HARGIS, G.K., WILLIAMS, G.A., REYNOLDS, W.A., CHERTOW, B.S., KUKREJA, S.C., BOWSER, E.W. and HENDERSON, W.J.: Effect of somatostatin on parathyroid hormone and calcitonin secretion. Endocrinology 201: 745-750, 1978. ). Algunas células parafoliculares tiroideas contienen somatostatina y calcitonina (VAN NOORDEN y cols., 1977[43]VAN NOORDEN, S., POLAK, J.M. and PEARSE, A.G.E.: Single cellular origin of somatostatin and calcitonin in the rat thyroid gland. Histochemistry 53:243-247, 1977.). La liberación de somatostatina puede conducir a una disminución de la secreción de calcitonina por un mecanismo autocrino (GORDIN y cols., 1978[44]GORDIN, A., LAMBERG, B.-A., PELKONEN, R. and ALMQVIST, S.; Somatostatin inhibits the pentagastrininduced release of serum calcitonin in medullary carcinoma of the thyroid. Clin. Endocrinol. 8 ; 289-293, 1978. ; REICHLIN, 1983[45]REICHLIN, S.: Somatostatin. N. Engl. J . Med. 309: 1495-1501, 1556-1563, 1983. ).
La somatostatina altera la agregación plaquetaria y bloquea la leucocitosis en respuesta a endotoxinas (McCANN, 1982[46]McCANN, S.M.: Physiology and somatostatin. Ann. pharmacology of Rev. Pharmacol. LHRH and Toxicol. 22: 491-515, 1982. ). Induce la secreción en los mastocitos (THEOHARIDES y cols., 1981[47]THEOHARIDES, T.C., BETCHAKU, T. and DOUGLAS, W.W.: Somatostatin-induced histamine secretion in mast cells. Characterization of the effect. Eur. J . Pharmacol. 69: 127-137, 1981. ).
La somatostatina no modifica la actividad de renina plasmática en sujetos sanos ni en pacientes hipertensos con renina baja; sin embargo, reduce significativamente los valores de renina de pacientes con hipertensión esencial con renina alta o normal (ROSENTHAL y cols., 1978a[48]ROSENTHAL, J., RAPTIS, S., ZOUPAS, C. and ESCOBAR-JIMENEZ, F. : Hemodynamic and renin responses to somatostatin in essential hypertension. Metabolism 27: 1361-1363, 1978a. , 1978b[49]ROSENTHAL, J., RAPTIS, S., ZOUPAS, C. and ESCOBAR-JIMENEZ, F. ; Inhibition by somatostatin of renin, blood pressure, and cardiac and stroke index in essential hypertension. Cir. Res. 43 (suppl.21: I-69-I-76, 1978b.).
Está implicada en la diabetes no insulino-dependiente y en la úlcera duodenal. Existen tumores pancreáticos de las células D.
En lo que respecta a la SST-28, se han estudiado sus acciones de la sobre la secreción hipofisaria humana, habiéndose demostrado que este polipéptido suprimía la respuesta de la GH a la arginina (RODRIGUEZ-ARNAO y cols., 1981[50]RODRIGUEZ-ARNAO, M.D., GOMEZ-PAN, A., RAINBOW, S.J., WOODHEAD, S., COMARU-SCHALLY, A.M., SCHALLY, A.V., MEYERS, C.A. and COY, D.H. : Effects of prosomatostatin on growth hormone and prolactin
response to arginine in man. Lancet 1: 353- 356, 1981. ; ) y a la hipoglucemia insulínica (MILLAR y cols., 1983). La SST-28 parece ser más potente y parece ejercer un efecto más prolongado que la somatostatina-14 (RODRIGUEZ-ARNAO y cols., 1981; MILLAR y cols., 1983MILLAR y cols., 1983[51]MILLAR, R.P., KLAFF, L.J., BARRON, J.L., LEVITT, N.S. and LING, N .: Somatostatin-28 and somatostatin-14 suppresion of arginine-, insulin- and TRHstimulated GH and PRL secretion in man. Clin. Endocrinol. 18: 277-285, 1983. ; GOMEZ-PAN Y RODRIGUEZ ARNAO, 1983[52]GOMEZ-PAN, A. and RODRIGUEZ-ARNAO, M.D.: Somatostatin and growth hormone releasing factor: synthesis, location, metabolism and function. Clin. Endocrinol. Metab. 12: 469-507, 1983. ). Las acciones de la SST-28 sobre la liberación de TSH parecen ser cualitativa y cuantitativamente similares a las ejercidas por la SST-14.
LA SST. REGULACIÓN
Por mecanismos dependientes del calcio iónico (IVERSEN y cols., 1978 [53]IVERSEN, L.L., IVERSEN, S.D., BLOMM, F., DOUGLAS, C., BROWN, M. and VALE, W .: Calcium dependent release of somatostatin and neurotensin from rat brain in vitro. Nature 273: 161-163, 1978.), los gránulos secretores de las terminaciones nerviosas SST, liberan en el sistema vascular portahipofisario las moléculas de SST, tras el estímulo de sus somas neuronales en los núcleos de origen, Igualmente, en las células no nerviosas que también secretan dicha hormona, los mecanismos secretorios también son dependientes del calcio iónico y los mecanismos que lo modulan.

Figura 6. Regulación de la somatostatina.
La dopamina estimula la liberación de somatostatina en fragmentos hipotalámicos de rata (MAEDA y FROHMAN, 1980) e incrementa la tasa de secreción y la concentración de somatostatina en la sangre porta hipofisaria cuando se administra a ratas por vía intravenosa (CHIHARA y cols., 1979; KATO y cols., 1979). Este efecto estimulante de la dopamina lo presenta también su precursor, la L-dopa, y la bromocriptina, un agonista de los receptores dopaminérgicos.
El ácido gamma-aminobutírico (GABA) inhibe la liberación hipotalámica de somatostatina (GAMSE y cols., 1980) aunque no afecta la secreción del péptido en otros territorios del sistema nervioso (SAWYNOK y cols., 1982).
Algunos neuropéptidos pueden influir también positiva o negativamente en la liberación de somatostatina. La sustancia P y la neurotensina son dos neuropéptidos capaces de estimular la liberación de somatostatina del hipotálamo de rata in vitro (PIMSTONE y cols., 1979 [54]PIMSTONE, B.L., SHEPPARD, M. , SHAPIRO, B. , KRONHEIM, S., HUDSON, A., HENDRICKS, S. and WALIGORA, K .: Localization in and release of somatostatin from brain and gut. Fed. Proc. 38: 2330-2332, 1979.; SHEPPARD y cols., 1979[55]SHEPPARD, M.C., KRONHEIM, S. and PIMSTONE, B.L.: Stimulation by growth hormone of somatostatin release from the rat hypothalamus in vitro. Clin. Endocrinol. 9 : 583-586, 1979. ) y de incrementar los niveles de somatostatina en la sangre porta hipofisaria (SAITO y SAITO, 1980[56]SAITO, H. and SAITO, S.: Effects of substance P and neurotensin on somatostatin levels in rat portal plasma. Endocrinology 107: 1600-1605, 1980.). La bombesina incrementa los niveles de somatostatina en la sangre porta hipofisaria (ABE y cols., 1981[57]ABE, H., CHIHARA, K., MINAMITANI, N. , IWASAKI, I., CHIBA, T., MATSUKURA, S. and FUJITA, T.; Stimulation by bombesin of immunoreactive somatostatin release into rat hypophyseal portal blood/ Endocrinology 109: 229-234, 1981.), lo que explica la reducción de los niveles circulantes de GH que origina. El péptido intestinal vasoactivo (VIP), sin embargo, estimula la liberación de GH a través de una inhibición de la liberación hipotalámica de somatostatina.
La glucosa se comporta como un inhibidor de la secreción de somatostatina. Pero cuando ésta cae en sangre se produce una estimulación de la liberación de somatostatina hipotalámica. De modo similar, una inhibición del metabolismo de la glucosa con 2-desoxiD-glucosa estimula la liberación de la hormona de una manera dosis dependiente. La hipoglucemia insulínica origina un incremento en los niveles plasmáticos periféricos de somatostatina, si bien el origen del péptido es difícil de precisar (WASS y cols., 1980[58]WASS, J.A.H., PENMAN, E., MEDBAK, S., DAWSON, A.M., TSIULAKIS, D., MARKS, V., BESSER, G.M. and REES, L.H.: Immunoreactive somatostatin changes during insulin-induced hypoglycaemia and operative stress in man. Clin. Endocrinol. 12: 269-275, 1980.). La insulina, per se, se comporta también como un estimulador de la liberación hipotalámica de somatostatina. (BERELOWITZ y cols., 1982[59]BERELOWITZ, M . , DUDLAK, D. and FROHMAN, L.A.: Release of somatostatin-like immunoreactivity from incubated rat hypothalamus and cerebral cortex. Effects of glucose and glucoregulatory hormones. J. Clin. Invest. 69: 1293-1301, 1982. )
El glucagón en administración intraventricular origina un incremento de la concentración de somatostatina en la sangre porta hipofisaria (ABE y cols., 1978[60]ABE, H. , KATO, Y. , CHIBA, T., TAMINATO, T. and FUJITA, T. : Plasma immunoreactive somatostatin levels in rat hypophyseal portal blood: effect of glucagon administration. Life Sci. 23: 1647- 1654, 1978. ). Sin embargo, la administración de glucagón por vía intravenosa carece de efecto sobre la secreción de somatostatina.
Hay evidencias de que la GH sea capaz de inhibir su propia secreción mediante un sistema de retroalimentación positiva de asa corta ("short-loop positive feedback") que actúe sobre la liberación hipotalámica de somatostatina. La administración intraventricular de GH es capaz de estimular la liberación de somatostatina hipotalámica de una manera dosis dependiente (CHIHARA y cols., 1981[61]CHIHARA, K., MINAMITANI, N., KAJI, H., ARIMURA, A. and FUJITA, T.: Intraventricularly injected growth hormone stimulates somatostatin release into ray hypophyseal portal blood. Endocrinology 109: 2279-2281, 1981. ). La GH también puede afectar la secreción de somatostatina a través de las somatomedinas. Se ha comprobado que la somatomedina A (SHEPPARD y cols., 1979[62]SHEPPARD, M.C., KRONHEIM, S. and PIMSTONE, B.L.: Stimulation by growth hormone of somatostatin release from the rat hypothalamus in vitro. Clin. Endocrinol. 9 : 583-586, 1979) y la somatomedina C (BERELOWITZ y cols., 1981[63]BERELOWITZ, M., PERLOW, M.J., HOFFMAN,H.J.and FROHMAN, L.A.: The diurnal variation of immunoreactive thyrotropin-releasing hormone and somatostatin in the cerebrospinal fluid of the rhesus monkey. Endocrinology 109: 2102-2109, 1981) son capaces de provocar una liberación de somatostatina en el hipotálamo de rata.
En los islotes pancreáticos, otros estímulos para la secreción de somatostatina por las células D los constituyen la glucosa, algunos aminoácidos y ciertas hormonas gastrointestinales como la gastrina, colecistoquinina, secretina y polipéptido inhibidor gástrico (SCHUSDZIARRA y cols., 1978[64]SZECOWKA, J., SANDBERG, E. and EFENDIC, S.: The interaction of vasoactive intestinal polypeptide (VIP), glucose and arginine on the secretion of insulin, glucagon and somatostatin in the perfused rat pancreas. Diabetologia 19; 137-142, 1980. ). El péptido inhibidor vasoactivo, la sustancia P y la neurotensina también liberan somatostatina de los islotes pancreáticos (SAITO y SAITO, 1980[65]SAITO, H. and SAITO, S.: Effects of substance P and neurotensin on somatostatin levels in rat portal plasma. Endocrinology 107: 1600-1605, 1980. ). El glucagón es un potente estimulador de la liberación de somatostatina por los islotes (SCHAUDER y cols., 1977[66]SCHAUDER, P., McINTOSH, C. and ARENDS, J.: Somatostatin and insulin release from isolated rat pancreatic islets in response to D-glucose, L-leucine, alpha-ketoisocaproic acid or D glyceraldehyde -evidence for a regulatory role of adenosines’ 5 ‘-cyclic monophosphate. Biochem. Biophys. Res. Commun. 75: 360, 1977. ). Por otra parte, se han detectado incrementos en los niveles plasmáticos de somatostatina en sangre arterial y en vena porta tras ingestión de varios alimentos (CHAYVIALLE y cols., 1980[67]CHAYVIALLE, J.A., MIYATA, M., ‘RAYFORD, P.L. and THOMPSON, i.e.: Effects of test meal, intragastric nutrients, and intraduodenal bile on plasma concentrations of immunoreactive somatostatin and vasoactive intestinal peptide in dogs. Gastroenterology 79: 844-852, 1980.; SCHUSDZIARRA y cols., 1980[68]SCHUSDZIARRA, V., ZYZNAR, E., ROUILLER, D. , HARRIS, V. and UNGER, R.H.: Free somatostatin in the circulation: amounts and molecular sizes of somatostatin-like immunoreactivity in portal, aortic, and vena caval plasma of fasting and meal-stimulated dogs. Endocrinology 107: 1572-1576, 1980. ). Todos estos hallazgos sugieren que la secreción de somatostatina se incrementaría durante el periodo de elevación de la concentración de nutrientes y hormonas gastrointestinales y que este péptido participaría en la regulación tanto de los procesos abortivos como de la función endocrina del páncreas (SCHUSDZIARRA y cols., 1978[69]SCHUSDZIARRA, V., IPP, E. , HARRIS, V,, DOBBS, R.E, RASKIN, P., ORCI, L. and UNGER, R.M.: Studies of physiology and pathophysiology of pancreaticD cell. Metabolism 27: 1227-1232, 1978.).
![]()
 Views Today : 40
Views Today : 40 Views This Month : 1730
Views This Month : 1730 Views This Year : 15495
Views This Year : 15495 Total views : 301558
Total views : 301558